生物科学研究所 研究報告
2025 年 7 月 1 日
ゲンジボタルの地理的変異と地質学的事件の関連
井口豊(生物科学研究所)
Report of Laboratory of Biology
July 1, 2025
Geographical variation in the firefly Luciola cruciata in relation to geological events
Yutaka Iguchi
Laboratory of Biology
I would like to dedicate this website to the memory of Dr. Nobuyoshi Ohba, who passed away on January 31, 2020. His memoir was published as "Memoir of Dr. Nobuyoshi Ohba" by Japan Association for Fireflies Research (Zenkoku Hotaru Kenkyukai).
The Genji firefly Nipponoluciola cruciata (formerly named Luciola cruciata) are classified into the three ecological types, the fast-flash, slow-flash, and intermediate types, on the basis of the regression of male interflash intervals on air temperatures (Iguchi, 2001, 2010). This classification has also been supported by molecular biological studies (Yoshikawa et al., 2001; Suzuki et al., 2002; Hiyori et al., 2007, 2010). Iguchi (2010) suggests that first the slow-flash type and then the intermediate type evolved from an ancestral fast-flash type. The intermediate type ("the West Japan 3" DNA type) may be named "the Fossa magna type", because it is endemic to the central part of Japan called "Fossa magna". Iguchi (2001) suggests that the separation between the slow- and fast-flash types was caused by the uplift of the Kanto Mountains at 11Ma and that the separation between the fast-flash and intermediate types was caused by the uplift of the Hida Mountains at 3Ma. These uplifts were presumably related to the movement of the Pacific and Philippine plates. Geographic variations in morphology of this species have been recently found in central Japan (Iguchi, 2024). Males have a wider body shape in Western Japan than in Eastern Japan (Iguchi, 2025).
References
Hiyori Y, Mizuno T, Kusaoke H (2007) The influence of an intentionally introduced population on the genetic structure of a native population in the Genji-firefly Luciola cruciata. Zenkoku Hotaru Kenkyukai-shi (Proceedings of the Japan Association for Fireflies Research) 40: 25-27. [in Japanese]. PDF
Hiyori Y, Ohata Y, Kusaoke H, Iguchi Y and Mitsuishi T (2010) Identification of the native habitats of alien populations of the Genji firefly Luciola cruciata by DNA analysis. Zenkoku Hotaru Kenkyukai-shi (Proceedings of the Japan Association for Fireflies Research) 43: 27–32. [in Japanese]. PDF
Iguchi Y (2001) Pattern and process of geographical diversification of flashes in the firefly Luciola cruciata in the northern part of Yamanashi Prefecture. Zenkoku Hotaru Kenkyukai-shi (Proceedings of the Japan Association for Fireflies Research) 34: 10–12. [in Japanese]. PDF DOI: 10.5281/zenodo.11210400
Iguchi Y (2010) Temperature-dependent geographic variation in the flashes of the firefly Luciola cruciata (Coleoptera: Lampyridae). J Nat Hist 44: 861-867.PDF
Iguchi Y. (2024) Male Morphological Dimorphism in the Genji Firefly Nipponoluciola cruciata in Central Japan. Advances in Entomology, 12: 203-209. DOI: 10.4236/ae.2024.123016
Iguchi Y. (2025) Male Morphological Difference in the Genji Firefly Nipponoluciola cruciata between Eastern and Western Japan. Advances in Entomology, 13, 249-254. DOI: 10.4236/ae.2025.132016
Suzuki H, Sato Y, Ohba N. (2002) Gene diversity and geographic differentiation in mitochondrial DNA of the Genji firefly, Luciola cruciata (Coleoptera: Lampyridae). Mol Phylogenet Evol 22: 193-205.
Yoshikawa T, Ide K, Kubota Y, Nakamura Y, Takebe H, Kusaoke H. (2001) Intraspecific genetic variation and molecular phylogeny of Luciola cruciata (Coleoptera: Lampyridae)inferred from the mitochondrial ND5 gene sequences. Japanese Journal of Entomology (New Series) 4: 117-127. PDF
ゲンジボタルの3型とその由来:東日本,西日本,中間(フォッサマグナ)型
1. 長野県のゲンジボタル
ゲンジボタル Nipponoluciola cruciata (以前は Luciola cruciata)の発光周期には地理的変異があり,その周期に応じて,短周期型(西日本型, 2 秒型)と長周期型(東日本型, 4 秒型)に分けられてきたことは有名である(例えば,大場,1988)。中間型(3 秒型)が存在するらしいことも,かなり前から知られていたが,一般的には,発光周期は東西2型に分類されると考えられてきた。
近年の分子生物学的研究によって,発光周期の東西両型に対応する遺伝子型も判明してきた( 吉川ほか, 2001; Suzuki et al., 2002)。しかしながら,中間型と遺伝子型の対応がはっきりしなかったため,中間型の存在が疑問視される時期もあった。その後,筆者が発光周期と気温の関係を詳しく調べたところ,中間型は文字通り,東西両型の中間周期を示す存在として区別されることが分かってきた。その上,遺伝子型との対応で,中間型が西日本型のひとつのサブグループに相当することも判明した(Iguchi,2010)。
図 1 を見て欲しい。
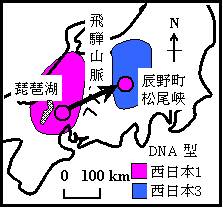
図 1. ゲンジボタル西日本型の2タイプの分布.長野県には西日本3型,関西には西日本1型が分布.ただし,関西からゲンジボタルを移入した辰野町松尾峡には西日本1型が分布.Iguchi (2009) より.
福井工業大学の草桶秀夫教授グループの研究によって,西日本型は飛騨山脈を挟んで東西に明瞭に異なる遺伝子型に分けられる(日和ほか,2007)西側の関西では 2 秒型,東側の長野県は 3 秒型となるのである(Iguchi,2010)。つまり,発光周期の観点から,一括して西日本型とはできないのである。飛騨山脈西側は「関西型」,東側(長野県)は「フォッサマグナ型」と呼ぶのが妥当であろう。
ここで,注意してほしいのは,2 秒,3 秒,4 秒というのは,気温 20℃での発光周期だということである。従来,気温を測定せずに,単純に周期のみを測定して,周期型を分類してきた。しかし,気温によって発光周期はかなり変化することが分かってきた(笹井; 1999; Iguchi, 2002; 翠川, 2006)。したがって,気温を考慮しないと地域間の比較は不可能なのである。これに関しては,松下(2021)が,全国各地のゲンジボタルに対して,気温と明滅周期の関係を DNA タイプと対比させて,精力的な研究を展開している。
発光周期の測定条件にも注意してほしい。ゲンジボタルのオスが交尾相手のメスを探す時,空中を漂うように飛翔 (hovering) しながら,集団同調 (mass synchronization) して発光するという特徴がある。この時の発光周期を測定し比較するのである。草木に止まっていたり,単独で飛んでいる個体の発光周期ではない。つまり,この種の特徴的な行動時の発光周期を測定するわけである。捕獲して室内に持ち込んで測定しても,野外の集団発光とは違う結果が出ることがあるので注意が必要である。
全国ホタル研究会誌では,20 ℃の時の発光周期を比較するべきという意見が出ていて(笹井, 1999; 翠川, 2006),私もこれに賛成である。
2. 天竜川上流,岡谷・辰野地域のゲンジボタルの生態的・遺伝的分析
1990 年代以後,私たち,岡谷市生物科学研究所や福井工業大学の草桶秀夫教授グループは,辰野町松尾峡だけでなく,その周辺地域のゲンジボタルの生態的・遺伝的調査を続けてきた。
下の図 2 のグラフ(Iguchi, 2002)を見て欲しい。天竜川沿いの岡谷市から辰野町にかけて,1990年代に調査研究された,ゲンジボタル明滅周期の気温に対する回帰分析および共分散分析(ANCOVA)の結果である(Iguchi, 2002)。
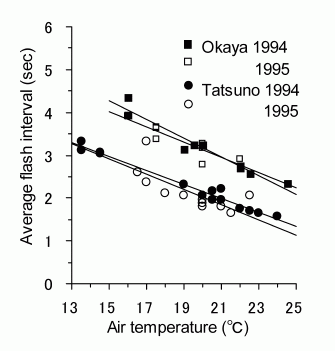
図 2. 長野県岡谷市駒沢と辰野町松尾峡における 1994,1995 年の気温とゲンジボタルの発光周期の関係.両個体群は天竜川沿いで 3km ほどしか離れていないが,関西からゲンジボタルを移入した松尾峡では短周期型となっている.Iguchi (2002) の図 1 より.
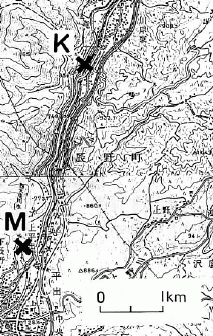
図 3. 天竜川沿いのホタル生息地.岡谷市 駒沢(K),辰野町 松尾峡(M). 井口(1993)図 1 より。
図 2 に示された岡谷市駒沢と辰野町松尾峡のゲンジボタルは,以前は,いずれも西日本型(2 秒型)とされてきた(三石,1990) 。しかし,このような回帰分析を行うと明瞭に発光周期が異なることが分かる。20 ℃で比較すると,岡谷市駒沢では中間型(3 秒型),辰野町松尾峡では西日本型(2 秒型)であることが分かってきた。実は,松尾峡のゲンジボタルは,人為的移入による外来種だったのである(辰野の移入(外来)ホタル 生物多様性の喪失へ)。
Yahoo!知恵袋で, 昭和30年代の天龍川水系で、特に岡谷辰野付近では元祖の蛍は生き残ることが出来ず、移入した在る種の蛍のみが生存できた
という仮説を提唱した人がいた(質問 生物の研究者とお見受けしますが
2015/5/27 23:53:06)
しかし,これは辰野町にとって都合よく解釈した誤解である。松尾峡,駒沢の天竜川沿いの個体群はわずか 3 kmほどしか離れておらず(図 3),ともに天竜川の水を取り入れた用水路に分布する。1960 年代ころ天竜川が汚れたからゲンジボタル個体数が減少したと言われるが,岡谷市駒沢のように生存し続けたゲンジボタル個体群も存在するのである。
2000 年代に入り,長野県内のゲンジボタル DNA の調査が進展した(図 4)。その結果,岡谷市 駒沢や辰野町 鴻ノ田の個体群は,フォッサマグナ地域固有の個体群であることが分かってきた(日和ほか, 2007)。この結果は,私の発光周期の研究結果(Iguchi, 2009)とも良く一致した。
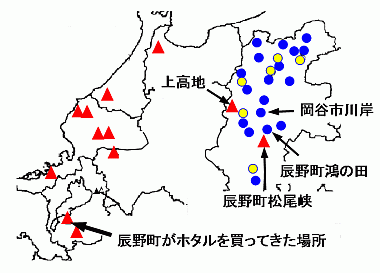
図 4. 長野県のゲンジボタルのDNA.辰野町松尾峡と上高地の個体群は関西から移入された外来種のDNAである。天竜川沿い,松尾峡の上流3kmの岡谷市駒沢の個体群は在来種のDNAである.この分布図は,日和ほか(2007)の図 1 に,なるべく忠実に基づいているので,実際の地図の地点とは多少ズレがある.
天竜川上流域のゲンジボタルの生息変化については,2015年10月10日に,滋賀県・守山市ほたるの森資料館で行なわれた私の講演会でも語られた(井口, 2015; 辰野のホタル 町おこしと保護の課題 - 滋賀県守山市・環境学習会)。
辰野町では,このような周辺地域のゲンジボタルの生息状況を把握することなく,観光優先で県外からの移入に頼ってしまった(井口, 2010)。辰野町誌の p. 538 にも,松尾峡周辺地域のゲンジボタルを採集して増やすのは苦労だから,業者を通じて外来種ゲンジボタルを移入したことが記されている(勝野, 1989)。松尾峡が1926年に県天然記念物に指定される際の調査でも,成虫発生期間後は,メスを安全な場所に移住させ卵を保護する
という提案が,県から示されていた(長野県, 1925)。しかし,実際にそのようにして松尾峡ゲンジボタルを増やそうという試みも,辰野町では実行されなかった。
このようにして,ホタルの名所辰野町松尾峡
では,外来種ゲンジボタルの移入という安易な行為によって,貴重な在来ゲンジボタルを保護することなく滅ぼしてしまった。現在,松尾峡で見られる多数のゲンジボタルは,この外来種個体群であり,そこに本来(県天然記念物指定時まで)生息していた個体群を増やしたものではない。
当時は,上述のような科学的データは得られなかった,という意見もあり,それには私も同意できる。しかし,現在見られる松尾峡ゲンジボタルが移入によって得られたものであるという現実は,皆が理解しておくべきだろう。そして,それが分かった現在ですら,残存する在来ゲンジボタルを,移入外来ゲンジボタルの影響から守ろうせず,調査すらしない,させない辰野町役場の現実があることも知ってほしい。
3. ゲンジボタル3型と地質学的イベントの関連性
ゲンジボタルのDNAと発光周期を対応させ,フォッサマグナ地域(特に南部)におけるその進化をまとめておく(Iguchi, 2010)。図 5 を見て欲しい。
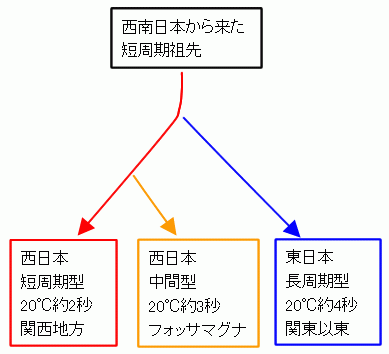
図 5. 中部地方におけるゲンジボタルの発光周期の進化。まず短周期型と長周期型が分岐し,次に短周期型から中間型が分岐したと考えられる。Iguchi (2010)より。
関西から関東にかけて,西日本由来の短周期型の祖先型から,まず東日本型が分岐し,その後,フォッサマグナ地域の中間型が分岐したらしい(Iguchi, 2010; 日和ほか,2010; 草桶ほか, 2011)。ただし,気温と発光周期の関係をさらに各地で調べれば,もっと複雑な分岐パターンが考えられるかもしれない。つまり,この3パターンだけでなく,もっといくつもの周期型に分かれている可能性がある。
長野・山梨両県の在来ゲンジボタル生息地を保護する人々は,生物多様性保全の観点からも,この貴重な「中間型」を守り,辰野町のようなことを起こさないでもらいたい。
今坂正一氏が,ゲンジボタル形態の地理的変異について,優れた研究を発表している(今坂,2010,2012,2013)。それによると,形態学的にも,フォッサマグナ東縁,つまり,関東山地付近で東西に分かれることが判明してきた。特に,雄の交尾器の差異は顕著であり,東西で別種あるいは別亜種となっている可能性も否定できない。私が形態データを多変量解析した結果も,関東山地付近で東西に分かれることを示唆する(Iguchi, 2024; 井口, 2025)。
筆者(井口, 2001a, 2001b; Iguchi, 2024;)は,中間型(フォッサマグナ型)と長周期型の分化は,関東山地の隆起に関連して約1100万年前に起き,一方,短周期型と中間型の分化は飛騨山脈の隆起に関連して約300 万年前に起きたと推定した。もしかすると,このような壮大なストーリーではなく,タイムスケールが1桁くらい異なるかもしれないが,ゲンジボタルの発光パターン,遺伝子,形態の地理的変異には,中部山岳地帯の地形形成が影響している可能性は大きい。そして,これらの山脈の形成史を考える時には,究極的にはプレートテクトニクス,つまり太平洋プレートやフィリピン海プレートの運動を考えることが不可欠となってくる。日本のような地質学的変動帯における生物地理学を考察するとき,プレートテクトニクスの議論は避けて通ることができない。このことは,伊豆諸島の生物地理とプレートテクトニクスでも触れた通りである。
以前は,フォッサマグナ西縁に当たる糸魚川-静岡構造線で,ゲンジボタルの発光周期は東西の型に分かれる,と思われたこともあった(大場,1988)。しかしその後,上述のように,発光周期は気温によって顕著に変化することが分かってきた。例えば,山梨県・下部は,従来は,東日本型(4 秒型)とされていたが(大場,1988),気温を考慮すると,中間型であることが分かり(Iguchi, 2010),遺伝子型の研究もそれを支持している(Suzuki et al., 2002)。
また,辰野町・松尾峡のゲンジボタルが西日本型(2 秒型)であるため,辰野町のやや東を通過する糸魚川-静岡構造線が東西 2 型の境界とされる理由でもあった(大場,1988)。しかし実は,松尾峡のゲンジボタルは県外から観光用に移入された外来種であり,大場氏が辰野町を訪問した時は,それが移入されたものだと役場から説明されなかったのである。
このような知見から,現在では,糸魚川-静岡構造線が発光周期の東西境界であるという考えは否定されている。
実際に,最近になって,長野県南西部の木曽地域,上松町から,辰野町 鴻ノ田と良く似た発光パターンを示す,中間型のゲンジボタルが発見されている(井口,2019,2020)。
最近では,群馬県と山梨県の間で,形態的な二型が生じていることが分かってきた。
4. 岡谷市における糸魚川-静岡構造線
岡谷市を含む諏訪盆地を通る糸魚川-静岡構造線は,国内第一級の活断層である牛伏寺断層の南延長であると考えられる(産業技術総合研究所・活断層・地震研究センター,活断層データベース 参照)。2014年には,この構造線北部の神城断層で地震が発生し,小谷村や白馬村を中心とした地域に大きな被害が生じた(長野県神城断層地震)。20112年に,岡谷市においても,この構造線に属する断層が新たに発見された(岡谷市で発見された糸魚川-静岡線に関連する活断層:岡谷市役所-敬念寺断層)。
さらに,同構造線に関連すると思われる活断層上にある岡谷塩嶺病院の敷地には,新たに看護学校が建設された(岡谷市看護専門学校(旧塩嶺病院)直下の活断層)。この活断層に関しては,岡谷市議会でも質疑・応答があったが,調査は行われずに,岡谷市看護専門学校として開校した。
なお,神城断層地震によって白馬村に現われた活断層の一部は,2015年に「神城断層」として白馬村の天然記念物に指定された(井口,2017c; 白馬村の天然記念物「神城断層」:2014年長野県神城断層地震の痕跡)。
図 6. 岡谷小学校跡地の位置。諏訪盆地および糸魚川-静岡構造線を一望できる位置にある。国土地理院 電子国土 Web システムによって表示されている。

図 7. 岡谷小学校の裏山から見た糸魚川-静岡構造線。上の地図のO地点から東方(地図上では右方向)を臨んだ風景。赤線が糸魚川-静岡構造線。諏訪盆地では,同構造線は複数の断層から成る断層群として表されるが,そのうちで古くから有名な断層を代表として表してある。
この写真(図 7)に示された糸魚川-静岡構造線(赤線)の東側(写真左側,諏訪盆地)と西側(写真右側,松尾峡)で,ゲンジボタル発光周期が異なるので,大場(1988)は,この構造線がゲンジボタル東西 2 型の境界と考えたのである。前述のとおり,現在では,そうでないと判明している。
図 6 に示された諏訪湖南西岸を通る断層は,かつては糸魚川-静岡構造線の主断層と考えられた(諏訪教育会,1975)。しかし現在では,諏訪湖南岸断層群と呼ばれ(澤ほか,2007),副次的な断層であり,東傾斜の撓曲構造の一部を成していることが分ってきた(池田ほか,2008: 文部科学省研究開発局ほか,2008: 澤ほか,2007;)
私は,岡谷市西部の山麓の断層の特徴を調査し,日本活断層学会2013年度秋季学術大会(つくば国際会議場)で発表した(井口,2013)。
その結果を見ると,断層の走向は,岡谷市やまびこ公園以北では,NS から NE-SW であり,以南では WNW-ESN であった。前者は中央隆起帯の伸び方向に一致し,後者は諏訪湖南西岸の断層の走向と調和的であった。しかも,ほとんどの断層が正断層であると推定されたが,調査地の最北部では逆断層とも見られる断層が発見された。岡谷市付近で,糸魚川-静岡構造線の向きや性質に変化があるのかもしれない。
塩嶺地域は,従来,塩尻峠ギャップと呼ばれ,糸魚川-静岡構造線の中でも,活断層が存在しない区間とされてきたが,最近になって活断層の存在が明らかになってきた(岡谷市看護専門学校(旧塩嶺病院)直下の活断層)。私の調査でも,塩嶺の看護専門学校直下を通ると思われる断層が確認された(井口,2013)。
5. 追悼
日本のホタル研究をリードしてこられた大場信義氏が,2020年1月31日に逝去された。全国ホタル研究会情報交換誌・第43号が,大場信義博士追悼号となって公開されている。
参考文献
日和佳政・水野剛志・草桶秀夫(2007) 人工移入によるゲンジボタルの地域個体群の遺伝的構造への影響. 全国ホタル研究会誌 40: 25-27.
日和佳政・大畑優紀子・草桶秀夫・井口豊・三石 暉弥(2010) 遺伝子解析による移植されたゲンジボタルの移植元判別法.全国ホタル研究会誌 43: 27-32.
井口豊 (1993) 長野県中部におけるゲンジボタルの発光パターン.月刊むし 263: 18-19. ResearchGate
井口豊(2001a) 山梨県北部におけるゲンジボタルの発光パターンと地理的分化の過程.全国ホタル研究会誌 34: 10-12. DOI: 10.5281/zenodo.11210400
井口豊(2001b) ゲンジボタルの明滅周期一気温関係の地理的変異と進化.日本鞘翅学会第14回大会講演要旨,p. 15.
Iguchi Y (2002) The influence of temperature on flash interval in the Genji-firefly Luciola cruciata (Coleoptera: Lampyridae). Ent Rev Japan 57: 119-122.
井口豊(2008) 中部地方におけるゲンジボタルの明滅周期について.全国ホタル研究会誌 41: 43-45.
Iguchi Y (2009) The ecological impact of an introduced population on a native population in the firefly Luciola cruciata (Coleoptera: Lampyridae). Biodiversity and Conservation 18: 2119-2126.
井口豊(2010) 長野県辰野町における移入ゲンジボタルについて.全国ホタル研究会誌 43: 23-26.
Iguchi Y (2010) Temperature-dependent geographic variation in the flashes of the firefly Luciola cruciata (Coleoptera: Lampyridae). J Nat Hist 44: 861-867.
井口豊 (2013) 長野県岡谷市の塩嶺西山地域における断層と地すべり地形. 日本活断層学会2013年度秋季学術大会講演予稿集: 60-61.
井口豊 (2015) ほたるの町 辰野(長野県)でのほたる育成の取組み.守山市ほたるの森資料館 2015年度 第1回環境学習会 講演: 2015年10月10日.
井口豊 (2017) 2014年長野県神城断層地震後の地形と植生の変化. 日本活断層学会2017年度秋季学術大会講演予稿集: 102-103.
井口豊 (2018a) 辰野の移入(外来)ホタル 生物多様性の喪失へ.生物科学研究所研究報告.
井口豊 (2018b) 辰野の在来ホタル:辰野に元々住んでいた自然のホタル.生物科学研究所研究報告.
井口豊 (2019) 上松町のゲンジボタルは,いつ,どこから来たのか? 信州環境カレッジ「上松町ホタル講習会」,長野県木曽郡上松町 下河原公園(2019 年 10 月 27 日). DOI: 10.5281/zenodo.12673464
井口豊 (2020) 長野県上松町のゲンジボタル.全国ホタル研究会誌 53: 23-24.
Iguchi Y. (2024) Male Morphological Dimorphism in the Genji Firefly Nipponoluciola cruciata in Central Japan. Advances in Entomology, 12: 203-209. DOI: 10.4236/ae.2024.123016
井口豊 (2025) ゲンジボタルの東西日本における形態的な違いについて. Authorea. DOI: 10.22541/au.174431375.56262712/v1. PDF
池田安隆・岩崎貴哉・伊藤谷生・狩野謙一・佐藤比呂志・阿部信太郎・金 幸隆・東中基倫・須田茂幸・川中 卓(2008) 糸魚川-静岡構造線中部を横切る反射法地震探査(辰野-諏訪側線).日本地球惑星科学連合 2008 年大会予稿集. S141-009.
今坂正一(2010) ゲンジボタルの地域変異について.第 1 報.鹿児島県~群馬県までの 12 地点の変異の計測結果.陸生ホタル生態研究会調査月報 24: 1-18.
今坂正一(2012) ゲンジボタルの地域変異について2.第 2 報.2010 年~2011 年に採集された宮崎県~群馬県までの 11 地点の計測結果.陸生ホタル生態研究会調査月報 39: 1-16.
今坂正一(2013) ゲンジボタルの地域変異について3.第3報.2012 年に採集されたヒガシゲンジボタル7地点の計測結果と今までのまとめ.陸生ホタル生態研究会調査月報 48: 1-27.
勝野重美(1989) 辰野のホタル.辰野町誌,570-612. 辰野町教育委員会.
草桶秀夫・木村和裕・日和佳政 (2011) 遺伝子から見たゲンジボタルの生物多様性と人為的放流の問題点.全国ホタル研究会誌 44: 13-19.
長野県(1925) 長野県史跡名勝天然記念物調査報告書.第三輯.
松下保男(2021) ゲンジボタル明滅周期の地理的分布とミトコンドリア DNA 多型の関係.陸生ホタル生態研究会調査月報 115: 1-17.
翠川博之(2006) ゲンジボタルの発光周期について.全国ホタル研究会誌 39: 32-36.
三石暉弥(1990) ゲンジボタル.信濃毎日新聞社.
文部科学省研究開発局・国土交通省国土地理院・東京大学地震研究所(2008) 糸魚川-静岡構造線断層帯における重点的な調査観測 平成19年度 成果報告書.
大場信義(1988) ゲンジボタル.文一総合出版.
笹井昭一(1999) ゲンジボタル明滅周期と気温について.全国ホタル研究会誌 32: 22-25.
澤 祥・谷口 薫・廣内大助・松多信尚・内田主税・佐藤善輝・石黒聡士・田力正好・杉戸信彦・安藤俊人・隈元 崇・佐野滋樹・野澤竜二郎・坂上寛之・渡辺満久・鈴木康弘(2007) 糸魚川-静岡構造線活断層帯中部,松本盆地南部・塩尻峠および諏訪湖南岸断層群の変動地形の再検討.活断層研究 27: 169-190.
Suzuki H, Sato Y, Ohba N. (2002) Gene diversity and geographic differentiation in mitochondrial DNA of the Genji firefly, Luciola cruciata (Coleoptera: Lampyridae). Mol Phylogenet Evol 22: 193-205.
諏訪教育会(1975) 諏訪の自然誌・地質編
吉川貴浩・井出幸介・窪田康男・中村好宏・武部寛・草桶秀夫(2001) ミトコンドリアND5遺伝子の塩基配列から推定されたゲンジボタルの種内変異と分子系統.昆虫(ニューシリーズ) 4: 117-127.