入倉・三宅の地震動評価式を大槻のデータで残差分析
井口豊(生物科学研究所,長野県岡谷市)
最終更新:2024 年 7 月 1 日
Residual analysis of the Irikura-Miyake ground motion model fit to Otsuki's data
Yutaka Iguchi (Laboratory of Biology, Okaya, Nagano, Japan)
Last Updated: July 1, 2024
PDF バージョン
DOI: 10.5281/zenodo.12606383
Abstract
Residual analysis is frequently used to evaluate the validity of statistical models. This article carried out the residual analysis of the Irikura-Miyake's (2001) ground motion model, log S (fault area) ∝ 1/2 log Mo (seismic moment) for the range between 7.5×1018 and 1.8×2018 (N·m), by using Otsuki (2016) compiled data. The results showed that the regression slope (that is, the scaling exponent) was expressed by 2/3 rather than 1/2. The residuals showed the possibility of a trimodal frequency distribution. The Irikura-Miyake model does not seem well-behaved for Otsuki's data and may be decomposed into more than two statistical models.
References
Irikura, K. and Miyake, H. (2001) Prediction of strong ground motions for scenario earthquakes. Journal of Geography 110: 849–875.
Otsuki, K. (2016) Kunihiko Shimazaki's comments to Nuclear Regulation Authority. Facebook, August 12, 2016. https://www.facebook.com/kenshiro.otsuki/posts/853391561458661?pnref=story (accessed September 8, 2016).
1. 入倉・三宅(2001)式とは
地震の規模(地震モーメント)と震源断層の長さまたは面積との関係(スケーリング関係)を記述する経験式として有名なものに,入倉・三宅(2001)の式がある。これについては,地震調査研究推進本部事務局の強震動予測手法,いわゆるレシピ
でも取り上げられている(地震調査研究推進本部事務局,2016)。例えば,その p.5 式 (3)' に記述された震源断層の面積 S (km2) と地震モーメント M0 (N·m) の入倉・三宅(2001)式は,以下のようになる。
要するに,断層の面積 S は地震モーメント M0 の 1/2 乗に比例するという,べき乗則(power law)
が,上記範囲において成り立つことになる。この 1/2 の部分はスケーリング指数(scaling exponent)とも呼ばれる。
これは,両対数座標でデータ点をプロットすると,ほぼ直線関係となることを意味する。ただし,元のデータに対して非線形回帰を行なってベキ関数を当てはめた場合と,対数変換して線形回帰によってパラメータを求め,改めてベキ関数に変換した場合は,式が異なってくる。ところが,これを知らない研究者もいるから注意が必要である。それについては,次のページ参照,Excelグラフ累乗,指数,多項式近似の論文記載の注意。
兵庫県南部地震(阪神・淡路大震災)が起きた1995年に,岩波書店の雑誌「科学」に掲載された島崎・長浜(1995)の論文は,地震の規模と断層の大きさのスケーリング関係をフラクタル理論と絡めて分かりやすく解説している。
2. 昆虫,特に甲虫のスケーリング関係
話は地震学から外れるが,生物学で,べき乗則と言えば,アロメトリー (allometry)という用語が,すぐに思い起こされる。しかし最近では,これもスケーリング関係(scaling relationship)と呼ばれることが多くなった(例えば,Emlen et al., 2007; Iguchi, 2013)。
私が研究しているカブトムシ(Trypoxylus dichotomus septentrionalis)では,興味深いことに,幼虫時代の栄養条件によって,成虫の体のサイズと角のサイズのスケーリング関係が異なる式で表されることが分かってきた(図1,Iguchi, 1998)。
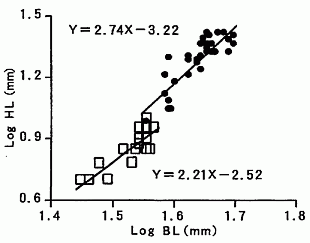
一方で,カブトムシやワガタムシの体のサイズと,それらの角や大顎のサイズは,単純なベキ関数よりも,ロジスティック曲線(logistic curve)と呼ばれる S 字曲線によって,しばしば表現されることも分かってきた。例えば,ダイコクコガネ Copris ochus のように,一見すると,直線的に見えるスケーリング関係であっても,統計学的に残差分析(residual analysis)をすると,ロジスティック回帰曲線が妥当であることが示されている(井口, 2014)。
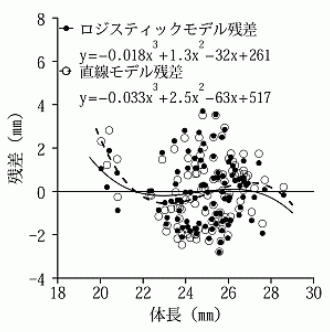
3. 入倉・三宅(2001)式の残差分析
入倉・三宅(2001)の式へ話を戻すと,最近では,福井県にある大飯原子力発電所の耐震設計基準をめぐる訴訟で,この式が一般の人々にも知られるようになった。これに関して,島崎邦彦氏が同式が地震モーメントを過小評価する傾向にあると主張し ており,私も日本活断層学会2015年大会(岡山大学)での島崎氏の発表(島崎,2015)を聞き,データ解析の観点から非常に興味を覚えた。
入倉・三宅(2001)式の評価を巡っては,大槻憲四郎(2016)が,Facebook 上で様々な観点から論じており,それも興味深い。今回,彼がコンパイルした地震モーメントと断層面積の関係を表すグラフ(図 3)を利用して,入倉・三宅(2001)式の残差分析を行ってみた。
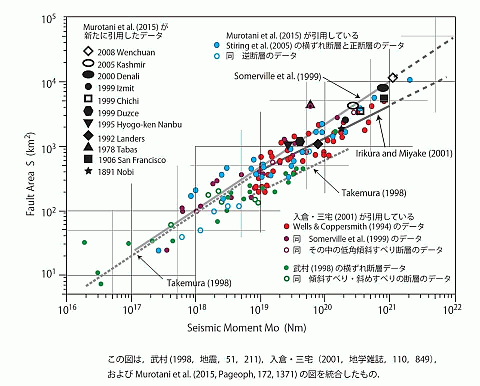
このグラフから,地震モーメント(N·m)が前述の範囲 (7.5×1018, 7.5×1018)にあるデータの残差分析を行なった。数値データは,北海道大学原子核反応データ研究開発センターが開発したグラフ数値読取システム(GSYS)を使い読み取られた。 その上で,地震モーメントに対する断層面積の残差変動,および,残差の頻度分布を調べた。読み取り誤差は含まれるものの,残差の特徴は捉えられると思われる。データ解析および作図には,統計解析ソフト R が用いられた。
4. 結果および考察
図 4 に,残差分析の結果を示す。横軸に地震モーメント,縦軸に入倉・三宅(2001)式からの断層面積のズレを取ってある。
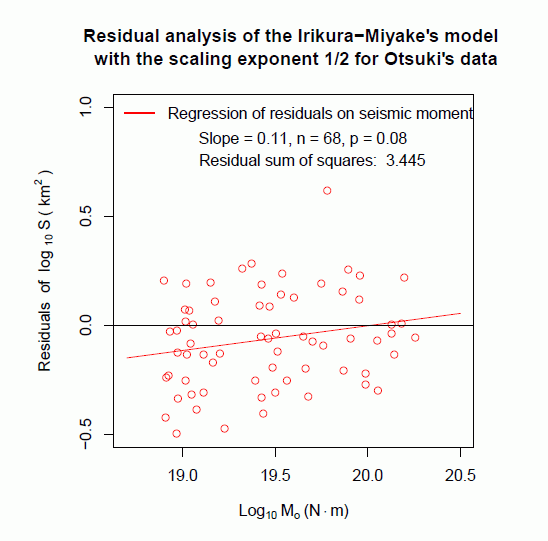
前述のダイコクコガネのスケーリング関係のように残差データの変動が波打っていないので,地震モーメントと断層面積の対数データが直線関係で表されることは,ほぼ確かなようである。したがって,この場合,べき乗則というよりも,両対数変換して直線関係で表される規則と呼ぶべきだろう。
一方で,もし残差が 入倉・三宅(2001)式を中心にして,上下にランダムに現われていれば,残差データの直線回帰は,傾き 0,切片 0 となるはずである。傾き 0 に対する検定結果は,p = 0.08 (df = 66) であり,やや疑問が残る,すなわち,スケーリング指数が 1/2 ではない可能性を示唆するものであった。
傾きと切片の値を固定せずに,回帰分析を行うと,地震モーメントと断層面積の対数データは,傾き(=スケーリング指数) 0.61 となり,それは 2/3 に近かった。そこで改めて,スケーリング指数を 2/3 に固定して log Mo に対する log S の回帰直線を求め,残差分析を行ってみた。その結果を図 5 に示す。
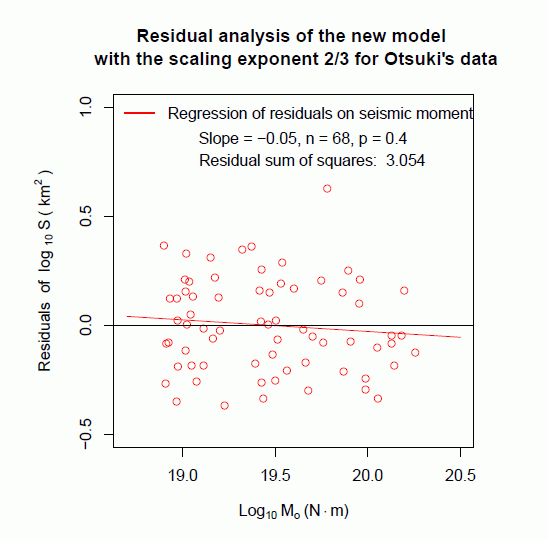
スケーリング指数を, 入倉・三宅(2001)式の 1/2 とした場合より,新たなモデルの 2/3 とした場合の方が適合度が改善され,残差平方和も減少した。
次に,入倉・三宅(2001)式の残差の頻度分布を調べてみると,以下の図 6 のようになった。
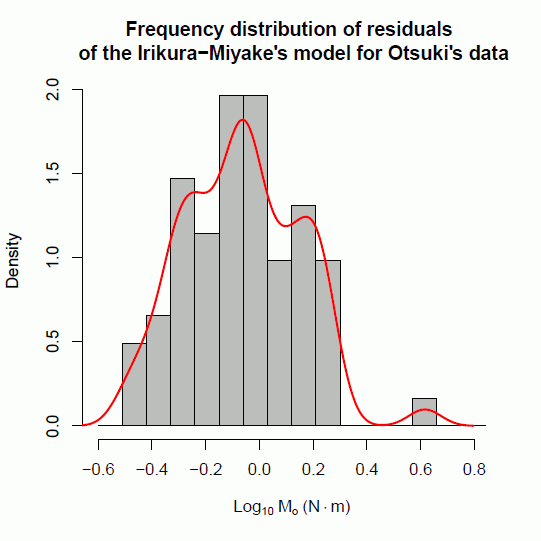
この図には,残差ヒストグラムに加え,カーネル密度関数で推定される曲線が描かれている。ここでは,ガウス型カーネル密度関数(Gaussian kernel density function)がバンド幅 0.06 で計算されている。
入倉・三宅(2001)式を適合させた場合の残差の頻度分布は,単一分布(unimodal distribution)でなく,その式の上下に二つの山を持ち,全体として三山分布(trimodal distribution)となった。
今回, 入倉・三宅(2001)式の適合が良くなかった理由は,いくつか考えられる。ひとつには,元データが適切に算出されていない可能性がある。例えば,入倉ほか(2015)は,武村(1998)の断層データは震源断層でなく地表断層を評価している,と指摘している。もうひとつには,大槻(2016)が指摘しているように,条件によって複数のモデルを使い分ける,複合モデルを考える必要があるかもしれない,ということである。この場合,もし単一モデルで代表させるとすれば,誤差を通常よりかなり大きく考える必要がある。
岩波書店の雑誌「科学」に掲載された島崎(2016)の論文に対しても,入倉(2016)は,入倉・三宅(2001)式が適合することを繰り返し述べているが,残差分析は行われていない。私個人としては,入倉・三宅(2001)式に対して,データの残差がどのように振舞うのか,それを見たいのだが,そのような分析は,なぜか行なわれないようである。
5. 大槻憲四郎先生のこと
余談だが,大槻先生は,私の学生時代の恩師であり,大学卒論でも大変お世話になった。大槻先生のことで,繰り返し思い起こすことは,「文献に頼るな,自分の頭で考えろ」と何度か言われたことである。正直言って,室内授業は,あまり覚えていない(と言うと怒られそうだ)。よく覚えているのは,構造地質学の野外実習で,牡鹿半島へ行き,断層や褶曲を観察したときのことである。「百聞は一見に如かず」とは,まさにこのことだと思った。現在,牡鹿半島の褶曲構造は,日本の地質構造100選(No.055 牡鹿半島の褶曲とスレート)にも選ばれている。
文献に頼らず,実地で調べてみよう,という考えは,ゲンジボタル(Nipponoluciola cruciata)の地理的変異を解明する際にも大いに役立った。それ以前は,大場(1988)により,糸魚川-静岡構造線で東西の二型に分かれるという考え方が有力だった。しかし実際にフィールドで調べて見ると,フォッサマグナ地域全体に中間型が分布しており,糸魚川-静岡構造線を挟んで変異するという現象は見られなかった(井口,2008; Iguchi,2010)。さらに,中部地方でこれら三型に分化した原因として,関東山地と飛騨山脈の隆起が関わっているらしいことも推測された(井口,2001a, 2001b)。
大槻先生のデータを残差分析しながら,改めて,「文献に頼るな,自分の頭で考えろ」という警句を思い出すのであった。
参考文献
Emlen, D. J., Lavine, L. C. and Ewen-Campen, B. (2007) On the origin and evolutionary diversification of beetle horns. Proceedings of the National Academy of Sciences, 104(suppl 1): 8661-8668.
Iguchi, Y. (1998) Horn dimorphism of Allomyrina dichotoma septentrionalis (Coleoptera: Scarabaeidae) affected by larval nutrition. Annals of Entomology Society of America, 91: 845-847.
井口豊 (2001a) 山梨県北部におけるゲンジボタルの発光パターンと地理的分化の過程.全国ホタル研究会誌 34: 10-12.
井口豊 (2001b) ゲンジボタルの明滅周期一気温関係の地理的変異と進化.日本鞘翅学会第14回大会講演要旨,p. 15.
井口豊 (2008) 中部地方におけるゲンジボタルの明滅周期について.全国ホタル研究会誌 41: 43-45.
Iguchi, Y. (2010) Temperature-dependent geographic variation in the flashes of the firefly Luciola cruciata (Coleoptera: Lampyridae). Journal of Natural History 44: 861-867.
Iguchi, Y. (2013) Male mandible trimorphism in the stag beetle Dorcus rectus (Coleoptera: Lucanidae). European Journal of Entomology, 110: 159-163.
井口豊 (2014) ダイコクコガネの大きさに関する小林論文を読んで:回帰分析におけるモデル選択.鰓角通信,28: 33-35.
入倉孝次郎・三宅弘恵 (2001) シナリオ地震の強震動予測.地學雜誌,110(6): 849-875.
入倉孝次郎・宮腰研・釜江克宏 (2015) 強震動記録を用いた震源インバージョンに基づく国内の内陸地殻内地震の震源パラメータのスケーリング則の再検討.日本地震工学会論文集,15(7): 141-156.
入倉孝次郎 (2016) 岩波科学2016年7月号の島崎邦彦氏の「最大クラスではない日本海『最大クラス』の津波―過ちを糺さないままでは『想定外』の災害が再生産される」へのコメント.入倉孝次郎地震動研究所ウェブページ, http://www.kojiro-irikura.jp/pdf/iwanamikagaku-comment-Jul13-2016.pdf 7月13日版(2016年9月8日アクセス).
神戸秀彦 (2017) 新規制基準下での原発差止め訴訟の考察 (2): 高浜 3・4 号機大津地裁決定と同大阪高裁決定を中心として.法と政治 68(2): 161-307.
地震調査委員会 (2016) 震源断層を特定した地震の強震動予測手法(「レシピ」)(平成28年6月10日)
大場信義 (1988) ゲンジボタル.文一総合出版.
大槻憲四郎 (2016) 原子力規制委員会に対する島崎邦彦前委員長代理のコメント: (6) 私の考え その4. facebook 2016年8月12日13時53分投稿. https://www.facebook.com/kenshiro.otsuki/posts/853391561458661?pnref=story (2016年9月8日アクセス).
島崎邦彦・長浜裕幸 (1995) 地震はでたらめに起こっているのか?: 地震の集団的性質と個別的性質.科学,65(4): 241-244.
島崎邦彦 (2015) 活断層の長さから推定される地震モーメント: 日本海「最大」クラスの津波断層モデルについて.日本活断層学会2015年度秋季学術大会講演予稿集: 50-51.
島崎邦彦 (2016) 最大クラスではない日本海「最大クラス」の津波―過ちを糺さないままでは「想定外」の災害が再生産される.科学, 86(7): 653-660.
武村雅之 (1998) 日本列島における地殻内地震のスケーリング則―地震断層の影響および地震被害との関連―.地震 2, 51: 211-228.