生物科学研究所 研究報告
2024 年 12 月 6 日
カブトムシ雄の角長と闘争頻度について
井口豊*
*生物科学研究所,長野県岡谷市
Report of Laboratory of Biology
December 6, 2024
Differences in the frequency of fights between minor and major males in the horned beetle Trypoxylus dichotomus septentrionalis (Coleoptera: Scarabaeidae)
Yutaka Iguchi*
*Laboratory of Biology, Okaya, Japan
Abstract
In the Japanese horned beetle Trypoxylus dichotomus septentrionalis, males are dimorphic in horn size (Fig. 1) and often fight with other males for food or females. Iguchi (2001) showed that large males tended to show fighting behavior more frequently than small males (Fig. 2, Mann-Whitney U-test, p < 0.01). This result was also confirmed by Brunner–Munzel test in this review (p < 0.01). However, in some cases, small males also show fighting behavior as frequently as large males, and therefore the variance of the number of fights was larger in small males than in large males (Fig. 2, F-test, p < 0.01). As a result, the horn size of small males did not influence the frequency of fights (Fig. 2, Kendall rank correlation, τ = 0.354, p > 0.1). Small males of this species seem to adopt flexible tactics in fighting frequency as well as well-known alternative tactics such as sneak mating.
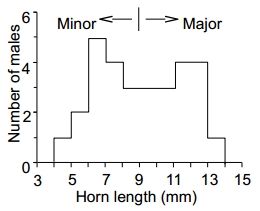
Fig. 1. Bimodal distribution of horn lengths (adopted from Iguchi 2001, Fig. 1).
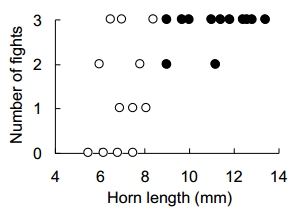
Fig. 2. Relationship between horn length and frequency of fights (adopted from Iguchi 2001, Fig. 3).
カブトムシの雄には,しばしば大型 major と小型 minor が存在し,日本のカブトムシ Trypoxylus dichotomus septentrionalis も,その角の長さの頻度分布を調べると,しばしば Fig. 1 のような二山分布 bimodal distribution となる。これを角の大きさに関する雄の二型という。
ただし,カブトムシ T. d. septentrionalis の場合,二型である(Iguchi, 1998)とも,三型である(Iguchi, 2000)とも断定しがたい面があり,今後,精査が必要だろう。私の研究(Iguchi, 2013)では,コクワガタ Dorcus rectus でさえ,二型でなく三型である可能性を示しているのだから,カブトムシの角やクワガタムシの大顎が二型であると安易に断定すべきではない。
さらに,一見すると,直線的に見える体と角のサイズの関係であっても,統計学的に分析すると,ロジスティック曲線(logistic curve)と呼ばれる S 字曲線の関係となっている場合もある。以下の Fig. 3 に示したダイコクコガネ Copris ochus のデータが,そのケースである(井口, 2014)。この分析では,赤池情報量規準 AIC (Akaike's Information Criterion) によって,ロジスティック曲線が判明した。
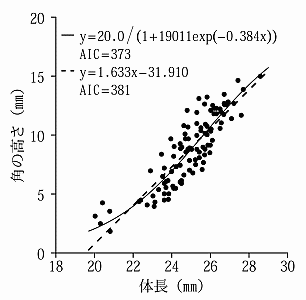
Fig. 3. Logistic curve fitted to horn size in Copris ochus. (adopted from Iguchi 2014, Fig. 1).
これらの甲虫の大型雄は闘争的であり,雄同士が戦うことによって,餌場や雌を獲得しようとする。一方で,小型雄は,むしろ戦いを避け,他の雄がいないときや,油断しているときを狙って,餌場や雌を獲得しようとする。これを代替的戦術 alternative tactics とか,代替的繁殖戦術 alternative reproductive tactics という。ただしこれは,カブトムシに限らず,クワガタムシ,さらには,動物一般に見られる行動である。
カブトムシの代替的戦術に関しては, Eberhard (1980, 1982), Emlen (1997), Hongo (2003), Karino and Niiyama (2006), Moczek and Emlen (2000), Siva-Jothy (1987) などが研究している。
Eberhard (1982) によれば, Podischnus agenor では,小型雄が大型雄より季節的に先に出現し,それが結果として,両者の間の闘争頻度を減じるのに役立っている。ただし私の研究で,日本のカブトムシを実験的に調べた結果では,大型雄のほうが小型雄より先に羽化するする傾向がある(Fig. 4)。日本のカブトムシでは,少なくとも出現時期に関しては,大きなサイズが大きな負担になっているとは言えないようである。
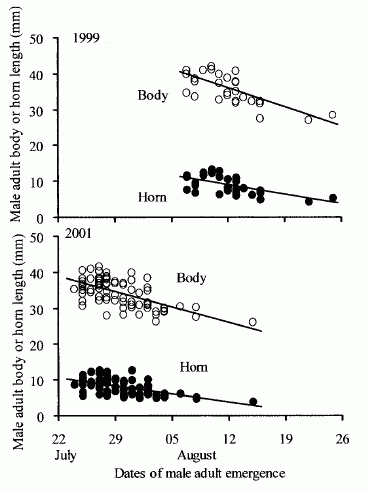
Fig. 4. Larger males emerge earlier in the season in the horned beetle Trypoxylus dichotomus septentrionalis. (adopted from Iguchi 2006, Fig. 1).
英語では,戦術 tactics の代わりに,戦略 strategy が使われることもある。辞書的なニュアンスから言えば,計画された「戦略」に基づいて,個々の「戦術」が実施されるのだろうが,カブトムシの行動研究では,これらの用語が,特に厳密に区別されているわけではない。
一方で,用語は厳密に区別せねばならぬ,という研究者もいて,例えば, Gross (1996) は, tactics と strategy をきちんと区別し,以下のように定義している。
A strategy is a genetically based program.
A tactic is a phenotype that results from a strategy.
やはり,これは辞書的な意味に似ている。
では,大型雄が小型雄より頻繁に闘争するとして,その頻度のバラツキはどうなのだろうか? つまり,ある大型雄の集団と小型雄の集団を比較した時,前者は,どの個体でも総じて闘争する頻度が高く,後者は,どの個体でも総じて闘争する頻度が低いと言えるだろうか?それを実験的に調べたのが, Iguchi (2001) である。
研究法は, Iguchi (2001) を参考にしてもらうとして,結果を Fig. 2 に示す。
確かに,大型雄のほうが,小型雄より闘争頻度が高かった(Mann-Whitney U-test, p < 0.01)。しかしながら,闘争頻度のバラツキの比(分散比)は,小型雄のほうが有意に大きかった(F-test, p < 0.01)。つまり,小型雄はどれでも闘争を避ける,というわけではなく,闘争的な個体もあれば,そうでない個体もある,というように,個体差が大きいのであった。
考えてみれば,当然の結果かもしれない。もし小型雄で,どの個体も総じて闘争的でなければ,角は文字通り,無用の長物 useless であり,それが無い個体が出てきても良さそうなものである。しかしながら,日本のカブトムシでは,どんなに小さな雄でも小さいながら角を持つ。小型雄の代替戦術は,闘争的ではないこと non-aggressive ではなく,闘争的であったりなかったりすること aggressive or nonaggressive であり,その戦術の柔軟性に小型雄の特徴があるのかもしれない。
なお本種の属名は,以前は Allomyrina であった。それを Trypoxylus に移した際には問題が生じた。以下のページを参照してほしい。
関連サイト
カブトムシ雌の闘争行動と同性間マウンティング
DOI: 10.5281/zenodo.14281281
References
Eberhard W.G. (1980) Horned beetles. Scientific American 242: 166-182.
Emlen D.J. (1997) Alternative reproductive tactics and male dimorphism in the horned beetle Onthophagus acuminatus (Coleoptera: Scarabaeidae). Behavioral Ecology and Sociobiology 41: 335-341.
Gross M.R. (1996) Alternative reproductive strategies and tactics: diversity within sexes. Trends in Ecology & Evolution 11: 92-98.
Hongo Y. (2003) Appraising behaviour during male-male interaction in the Japanese horned beetle Trypoxylus dichotomus septentrionalis (Kono). Behaviour 140: 501-517.
Iguchi Y. (1998) Horn dimorphism of Allomyrina dichotoma septentrionalis (Coleoptera: Scarabaeidae) affected by larval nutrition. Annals of the Entomological Society of America 91(6): 845-847.
Iguchi Y. (2000) Male trimorphism in the horned beetle Allomyrina dichotoma septentrionalis (Coleoptera, Scarabaeidae). Kogane 1: 21-23. (日本語タイトル:カブトムシのオスの3型について). ResearchGate.
Iguchi Y. (2001) Differences in the frequency of fights between minor and major males in the horned beetle Trypoxylus dichotomus septentrionalis (Coleoptera: Scarabaeidae). Entomological Review of Japan, 56 (1): 11-14.
Iguchi Y. (2006) Are beetle horns costly to produce? Evolutionary Ecology Research 8: 1129-1137.
Iguchi Y. (2013) Male mandible trimorphism in the stag beetle Dorcus rectus (Coleoptera: Lucanidae). European Journal of Entomology 110: 159-163.
井口豊 (2014) ダイコクコガネの大きさに関する小林論文を読んで:回帰分析におけるモデル選択. 鰓角通信, 28: 33-35.
Karino K. and Niiyama H. (2006) Males with short horns spent more time mating in the Japanese horned beetle Allomyrina dichotoma. acta ethologica 9: 95-98.
Moczek A.P. and Emlen D.J. (2000) Male horn dimorphism in the scarab beetle, Onthophagus taurus: do alternative reproductive tactics favor alternative phenotypes? Animal behaviour 59: 459-466.
Ratcliffe B.C. (2008) Book Review. Atlas of Japanese Scarabaeoidea. Volume 2. Phytophagous Group I. The Coleopterists Bulletin 62(1): 63-64.
Siva-Jothy M. T. (1987) Mate securing tactics and the cost of fighting in the Japanese horned beetle, Allomyrina dichotoma L. (Scarabaeidae). Journal of Ethology 5: 165-172.